"To understand it you need not only to create it, but also to break it"

"Yablokoshki (1)". Nadia Bazlova and Carlos Gershenson, St. Petersburg, 2000. Gel on paper.
In this chapter we will describe experiments carried out in our Behaviours Virtual Laboratory. In the first section, we will present experiments involving the emergent intelligence, opportunism, and lower cognition given by BeCA in the animats, while in the second section we will present experiments involving emergent social behaviours and structures in small animat societies. We encourage the readers to access our BVL through our web page https://bingweb.binghamton.edu/%7Ecgershensong/unamjlagunez/asia/bvl to execute the experiments presented here, and also to develop their own.
The goal of the experiments presented here is to show the properties and capabilities of BeCA, I&I, and our BVL.
In the first section, we will present experiments showing the simulation of reflex, reactive, and motivated behaviours. We will also show experiments involving primary and secondary classical conditionings, and the learning of the motivation degree. We will also present an experiment showing the non persistence of a consummatory behaviour in the presence of an aversive stimulus, and we will show experiments involving different motivation and reactive degrees through the adjustment of parameters in BeCA.
In the second section, we will first present simple experiments showing the imitation and induction of behaviours. Then we will expose the relevance of the environment in the sociality of the animats as they learn the imitation factors. Finally, we will present experiments concerning collective misbelief.
"Intelligence is the art of getting away with it"
--Arturo Frappé
We will present here experiments related with the adaptive intelligence emerging from BeCA, which provides the control for the animats.
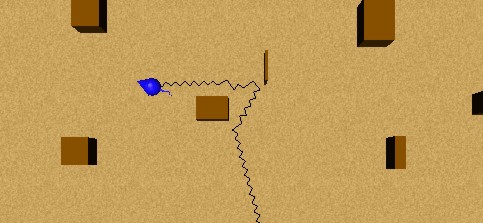
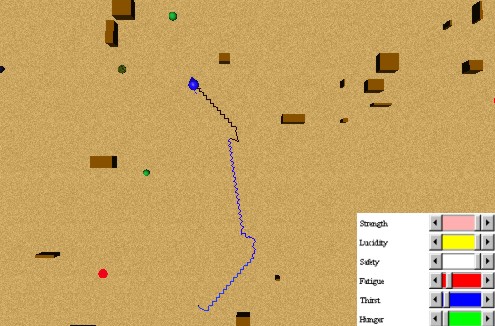
In this simple experiment, an animat is exploring following a straight trajectory. When an obstacle is perceived as too close, he reflexively avoids it. These behaviours can be appreciated in Figure 26. In animals, the behaviour "avoid obstacle" would be more reactive than reflex, but our animats have no planning, and this behaviour can then be seen as a reflex behaviour, because the signal to avoid the obstacle is inscribed on the External Perceptions level only when the obstacle is within a close range (2).
In this experiment we have a wandering prey animat and a hungry predator, as shown in Figure 27. But when the prey animat perceives the predator, he reactively runs away, as seen in Figure 28. We can see that the presence of the predator was enough to trigger the runaway behaviour, but not as directly as with reflex behaviours. If there are other stimuli that may trigger reactive behaviours, BeCA will select the most imperative one, depending on the external signals, to be executed (3).
In this experiment we have an initial state as the one shown in Figure 29. We have an animat with some hunger and a lot of thirst. He is perceiving a food source. Since the thirst column wins the competition at the motivational level (4), he does not approach the food, but begins to explore. Once he perceives a water source, he approaches it and drinks until his thirst is satiated. After this, the hunger column wins the competition at a motivational level, so he approaches the food and eats until he satiates his hunger. After this, he wanders because he has no internal needs, nor are there any stimuli that trigger reactive or reflex behaviours. This behaviours patterns can be appreciated in Figure 30.
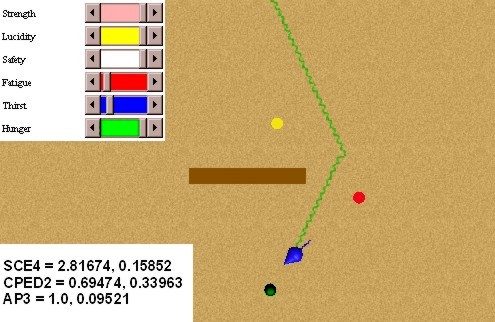
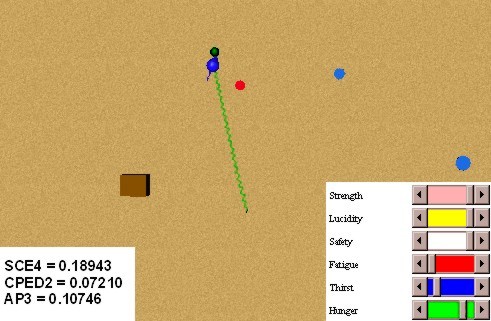
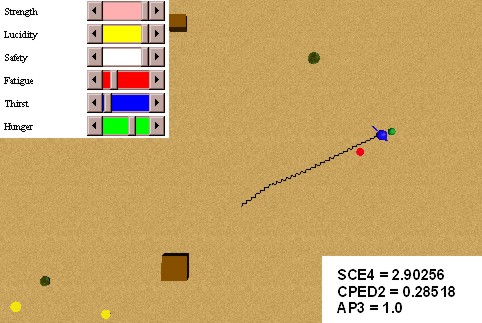
In this experiment we will test primary and secondary classical conditionings. In order to obtain the conditioning of a red spot with a food source, we make several presentations of the pair red spot - food source, when the animat is hungry. Figure 31 shows an initial stage of the conditioning: we can see that the coupling strengths related to the internal behaviours external behaviours selector (SCE4), intero/extero/drive congruence (CPED2), and attention to preferences (AP3), have still small values. Figure 32 shows an advanced stage of the conditioning, where the coupling strengths have higher values. The speed of the conditioning can be modulated in the BVL with the parameter lambda of expression (10) (5).
In Figure 33 we can see a behaviours pattern executed once the conditioning of the red spot with the food source was achieved. When the hungry animat perceived the red spot, he approached it, because the red spot was able to activate the same signal on the Actions level that a perceived food would have produced. That is to say, the modification of the coupling strengths allowed the red spot to complete a behavioural column, associated with the appetitive behaviour "approach food". Once the animat perceived a food source, he approached it, and satiated his hunger.
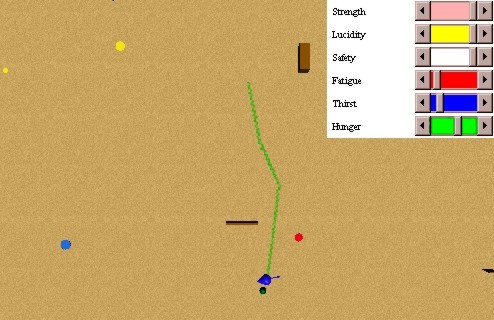
Once the red spot is conditioned with the food source, we can use the red spot as a conditioner of another neutral stimulus. In this case, yellow spots. In Figure 34 we can see an initial stage of the secondary conditioning (6). The values of the coupling strengths before the comma are the ones of red spots, while the others are the ones related with yellow spots. Before perceiving a red spot, the hungry animat perceives a yellow spot. If we repeat this situation, the coupling strengths will be reinforced, forming a behavioural column also for the yellow spot signal. In Figure 35 we can appreciate a behaviours pattern once the secondary conditioning was achieved. If the conditioned hungry animat perceives a yellow spot, he will approach to it, and once he perceives a food source, he will approach to this one, and thus satiate his hunger.
For this experiment we considered an initial state as the one shown in Figure 36. We have two animats in separated environments: one with abundant sources of food, and the other one with none. Other stimuli are created in both environments. Both animats are initialized with the same parameters: a very high level of hunger, and no other internal need, with an initial value of alfa for the hunger column of 0.7. alfamax has a value of 1.0, alfamin of 0.0, delta of 1000, and rho of 0.0005.
Figure 37 shows what occurred after 1000 simulation cycles. The animat in the scarce environment explored, searching for food, but finding none. This remembered scenario led to the incremental adjustment of his respective alfai until it reached alfamax. On the other hand, the abundant environment allowed the animat in it to satisfy his hunger quickly. Once his hunger was satiated, the animat wandered, while his respective alfai decreased (7).

We now can set two animats with the same degree of hunger, but with different alfai values. The animat with a high alfai, since he learned that food is scarce and will be difficult to find, will begin to explore. Meanwhile, the animat with a low alfai will wander, because he learned that he will find abundant food with little effort, and thus allows himself to wander, and will not explore until his hunger level is higher. An example of these behaviours can be seen in Figure 38.
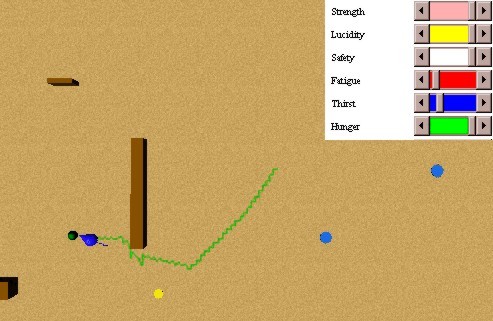
In this experiment we will test the non persistence of a consummatory action (eat) of a prey animat in the presence of a predator. We have an initial state as the one shown in Figure 39: a hungry prey animat, perceiving a food source, but not perceiving a thirsty predator that is nearby. The prey animat perceives the predator while he is eating, and runs away before he satiates his hunger completely, as seen in Figure 40.

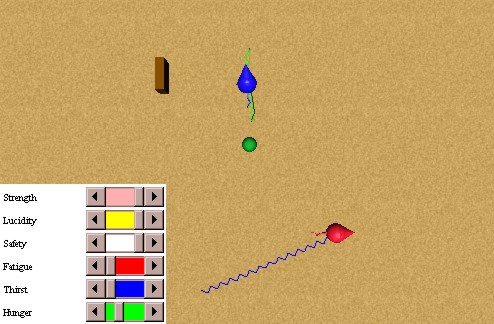
Afterwards, the prey animat perceived another food source, which he was able to satiate his thirst with, as seen in Figure 41. We used a value of kappa equal to 0.9. Otherwise, the persistence would have been enough to satiate the hunger of the prey animat. If the prey animat would eat slower, with a usual value of kappa we would have obtained similar results.
"We have to break things, but then we have to find out what to do with the pieces"
--Mafalda
In these experiments, we modified the values of the parameters alfa, gamma, and phi of BeCA; in order to observe how motivated or reactive is the behaviour of an animat depending on these parameters. Some of the adjustments of these parameters can be seen as lesions in different mechanisms of BeCA.
We used for all experiments an initial environmental state as the one shown in Figure 42. The animat has little fatigue, much thirst, and some hunger. There are food sources near him, but the water sources are distant and the animat cannot perceive them at this stage. The BVL's interface allows the easy loading of this initial state for each experiment.
First, we took values of alfa=0.8, gamma=0.0, and phi=1.0. These are the default values used in the BVL. The behaviours executed by the animat can be appreciated in Figure 43. Since the predominant need was thirst, the animat began to explore in the search of water, in spite of perceiving food sources. When he perceived a water source, he approached it, and drank until his thirst was satiated. After this, he approached a food source, and satiated his hunger by eating. These behaviours were motivated by the internal states of the animat.

For the next experiment, we took values of alfa=0.0, gamma=0.0, and phi=1.0. Since the internal and external signals are being combined multiplicatively (as it can be seen in expression (7)), the animat will need to perceive the water before the respective behavioural column may win the competition in the motivational node. So, since the animat was perceiving food and was hungry, he approached it and ate until his hunger was satiated. After this, he wandered until he finally perceived the water source, approached it, and satisfied his thirst by drinking it; as shown in Figure 44. These behaviours are less motivated, since the animat cannot execute the explore behaviour, and any internal need can fire its behaviour only if a corresponding external stimulus is perceived. This also affects the animat's survival performance, because he will need more time to find a stimulus to satiate a need than for higher values of alfa (González et. al., 2000).
For the following experiment, we used values of alfa=0.8, gamma=0.0, and phi=0.0. Since phi=0 causes that no signal from the motivational node reaches the cognitive node, the animat has no awareness of his internal needs. Because of this, he will wander independently of his perceptions or motivations, unable to perform motivated behaviours; as shown in Figure 45, until his death.
For values of alfa=0.0, gamma=0.0, and phi=0.0 we had similar results. If there is no flow of signals from the motivational node, the value of alfa will not affect the external behaviour of the animat.
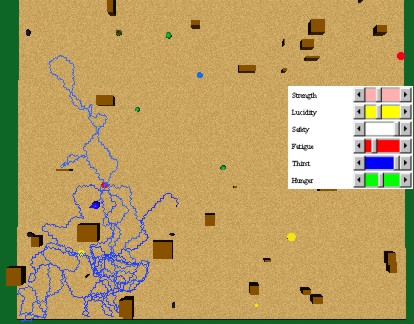
Next, we used values of alfa=0.8, gamma=0.1, and phi=1.0. gamma greater than zero in expression (4) allows a behaviour to be reactively executed, even in the absence of an internal need. Figure 46 shows the behaviours that the animat executed: First, he perceived food, approached it, and ate it completely, even when he had no more hunger. Then, he began to explore, until he perceived the water source, approached it, and drank it completely. Since he perceived a food source, he approached it in order to eat it reactively.
With values of alfa=0.0, gamma=0.1, and phi=1.0 we had the same results.
Figure 47 shows the last experiment, where we first took values of alfa=0.8, gamma=0.1, and phi=0.0. In this case, the animat has no awareness of his internal state, but his behaviour is reactive since gamma is greater than zero. He is able to reactively satiate his hunger and thirst, but because the external signals were at hand. Otherwise, he would have wandered independently of his internal needs. In this situation, the animat would only survive if he finds by chance external stimuli for the needs he has in a precise moment. If he does not run into an appropriate external stimulus, he would die unknowingly.
With values of alfa=0.0, gamma=0.1, and phi=0.0 we had similar results.
We can see that motivations are important for the survival of the animats. Although they could survive without them, being all the behaviours reactive, they have a higher probability to survive a scarce environment and adapt in it if their behaviours are motivated.
In this section we will present experiments involving small animat societies, and the complex social behaviour that emerges from their simple interactions, provided by our I&I model presented in Chapter 4.
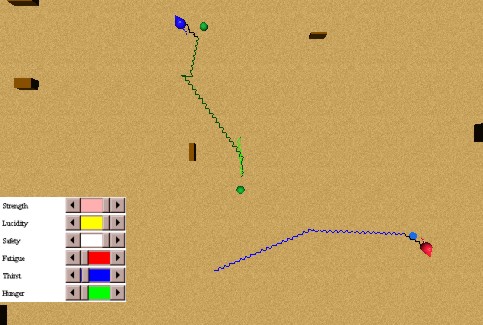
In this experiment we will test the imitation of a behaviour provided by our I&I model. Figure 48 shows the initial state of the experiment: a thirsty animat perceiving a water source, and a second thirsty animat perceiving the first, but without perceiving the water source. The first animat approaches the water source obeying his thirst. When the second animat perceives the first one approaching water, he imitates his behaviour because he is also thirsty and approaches him (as if the first would represent a water source). Once he perceives the water source, he approaches towards it directly. He avoids the first animat, who while drinking water does not allow the second animat to approach enough to the water source to be able to drink. When the first animat finally moves and the water source is free, the second animat satiates his thirst. These behaviours are illustrated in Figure 49.

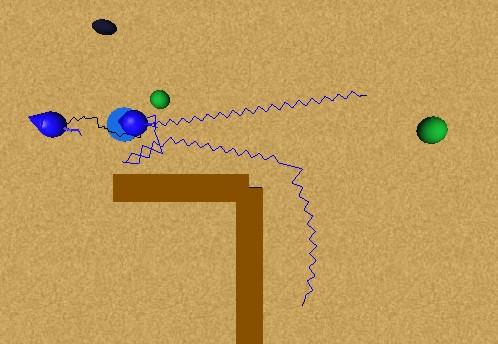
The imitation of behaviours increases the adaptiveness of the animats, and also improves their chances of survival. For higher values of lamed, the animats will be more social, and vice versa.
In this experiment, we will test the induction of behaviour, provided by I&I. We have for the initial state a predator animat, and two prey animats, as seen in Figure 50. One prey is perceiving the second prey and the predator, while the second prey is not perceiving the predator nor the first prey. When te simulation begins, the first prey animat begins to run away, but also induces the second prey to run away. He imitates the induced behaviour because of his need for safety, and runs away from the predator, as shown in Figure 51.


The induction of behaviours also increases the adaptiveness in animats. Animats with a higher value of yud will have a higher possibility to induce the rest of the animats. This may lead to the emergence of social hierarchical structures.
"Sometimes our intentions are not responsible for our possibilities"
--Nadia Bazlova
For these experiments, we created five prey animats with initial values of lamed equal to 0.5, lamedmax equal to 0.8, lamedmin equal to 0.2, and yud equal to 0.3.
First, we put the animats with some internal needs in a scarce environment, as seen in Figure 52. After more than ten thousand time cycles, half of the lamed values for imitation were in the neighbourhood of lamedmax, and half in the neighbourhood of lamedmin. This was because, in a scarce environment, animats tend to have similar internal needs, and thus imitation of behaviours can be performed, and lamed would be increased. We also made an experiment with the same initial state, only that one animat had a value of yud equal to 1.0. This did not make much difference in the adjustment of the lamed's, because other animats did not have a higher percentage of high values of lamed for the animat with a high yud than for others at the end of the experiment. The alfai values related with the internal needs that could not be satisfied were increased, increasing the motivation degree.
Next, we put the animats with no internal needs in an abundant environment, as shown in Figure 53. After more than ten thousand simulation cycles, all lamed's were below their initial value, and most in the neighbourhood of lamedmin. This is because, in an abundant environment, animats are wandering most of the time (remember that the adjustment of lamed's only takes place when the individual has internal motivations), and when one has internal needs enough to trigger an appetitive or consummatory behaviour, in most of the cases the animats he is perceiving are performing different actions, making all lamed's to tend to lamedmin. For this case we also made an experiment putting an animat with a value of yud equal to 1.0, but again this did not made a noticeable difference in the lamed values. Because of the abundant environment, the alfai values related to the internal needs related to the abundant stimuli decreased, decreasing the motivation degree.
A high value of yud does not make a big difference because the behaviours produced by BeCA were strongly motivated. For reactive behaviours, a high value of yud makes a more noticeable difference.


We could see by the previous experiments that a scarce environment makes the animats more social, while an abundant one makes them more selfish. Since the behaviour of the animats was motivated, this indicates that the environment shapes the society, more than its members do (if behaviours are motivated). This suggests sociology not to study only the individuals in order to study a society, but also their environment and context.
We also made experiments with an environment similar to the one shown in Figure 52, but with a non-homogeneous society of animats. We found that animats tend to be less social in such situations. Especially, reactive animats (gamma=0.1) tended to be less social than animats with a degree of motivation (alfa) equal to zero, and these also tended to be less social than animats with a high degree of motivation. Also, animats with a higher radius of perception (rp) tended to be less social than ones with a lower one. The perceptual persistence (kappa, aleph) seemed not to play an important role in the learning of the imitation parameters. Without taking into account sociality, animats with a higher degree of motivation (animats predisposed to be more social) have higher probability of survival , but animats with a higher radius of perception (animats predisposed to be less social) also have a higher probability of survival. This suggests that societies can tend to be more or less social (after some generations) depending on parameters that at first sight are not related with sociality. Of course, the environment also plays a very important role in the shaping of the society.
We could see that the shaping of a society is indeed complex, because it depends on so many elements: the behaviour of the individuals, their interactions, and their environment. A group can be more or less social because of several parallel reasons. Artificial societies and virtual laboratories are necessary tools in the understanding of such complex processes.
"All ideas are valid in the context they were created"
In these experiments we will study collective misbelief (8). An example of collective misbelief follows: Sven believes that Thorfin is going for wine, so he is going along with him; but Thorfin believes that Sven is going for wine, so he is going along with him. In fact, no one has wine or money to buy wine, but they have misbeliefs that are reinforced mutually. So, we can say that collective misbelief is given when a group of individuals believes that the rest of the group is knowing what they are doing, and vice versa, when actually no one knows what to do. Quoting Immanuel Kant, "one milks the male and the other one holds the bucket".
In order to simulate this, we thought of a configuration as the one shown in Figure 54: we have four hungry predator animats in a closed space with only one exit, each one perceiving another, and the last perceiving the first. Using clock coordinates, the first animat is at nine o'clock, the second at six o'clock, the third at three o'clock, and the fourth at twelve o'clock. The first predator is perceiving a prey on the entrance of the corral where the predators are almost confined to. We used values of rp equal to 30.0, aleph equal to 0.5, and yud equal to 0.0 to avoid induction of behaviour (otherwise, the first animat would tell the fourth where the prey was, and the fourth the third, and so on).

When the simulation starts, the first predator perceives the prey, and he begins to approach him. Since the second animat is also hungry, and perceives the first one approaching a prey, he imitates his behaviour and approaches him. The same occurs with the third perceiving the second, and the fourth the third. But, soon after the prey leaves the sight of the first animat, he forgets where the prey was, because of his value of aleph. So, since he is perceiving the fourth and second animats approaching to a prey, he imitates their behaviour, and approaches the second animat. So, what occurred was that the first approached the second and vice versa, and the third approached the fourth and vice versa, as seen in Figure 55. When they collided, they avoided each other, so the misbelief was broken, and everyone began to explore, as shown by Figure 56.

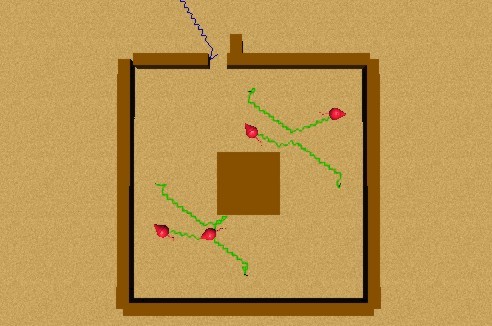
We thought about an experiment where the collective misbelief would not be broken, or at least not as easily as in the experiments just presented above, but none of our attempts were successful. Surprisingly, the situation was given by itself, when we were performing an experiment on the modification of the lamed's in a scarce environment. This unexpected result (as others we have had, such as the emergence of a "nesting" behaviour) was possible because of the complexity of the BVL and its components: BeCA and I&I.
We observed the behaviours and internal variables of the animats, and concluded the following: At least one animat conditioned a red spot with grass. He had fatigue, so he approached a red spot, but without any grass nearby. Without sociality, there would be an extinction of the conditioning, but another animat with fatigue perceived the first one approaching grass (the red spot), and he approached him. The fact that both animats were perceiving each other approaching grass, made them, not only not to lose the conditioning, but even reinforce it. Another animat with fatigue could fall also in the misbelief. When they collided with each other, they moved away from the red spot, but since they remembered its location, they returned again and again. So, they were moving around the red spot, with the imitation and induction of the other animats keeping them in the area, as seen in Figure 57. We then put two more animats with fatigue in the vicinity, and they easily fell in the misbelief, as seen in Figure 58.


Believing emerges from the imitation of behaviour (i.e. we can perceive beliefs in the behaviours of the animats, but there is no "belief module" containing them). This is, if one animat imitates another, it is because he believes in the correct behaviour of the imitated animat. Of course, as we could just see, beliefs are not always correct. They should not be always correct, which is why they are beliefs.
"There are reasons of the heart, which the heart does not know"
--Blaise Pascal
As we could see in the this chapter, animats in their virtual environment behave as animals in a natural environment. Of course the analogy is not total, but the main characteristics of adaptive animal behaviour were able to be reproduced in the BVL, because of BeCA and I&I.
We say that intelligence emerges in the animats because we, as external observers, can judge that their behaviour is intelligent because by adapting to their environment they are able to survive (not always, of course), but they take every opportunity to achieve it. This is for us intelligence. We say that the animats have emergent cognition, because for external observers, they know their environment, and what to do in it in order to survive.
We say that social phenomena and structures emerge in animat societies because we can perceive social behaviours, such as leadership (given by a high value of yud), organization (when animats perform the same behaviour to obtain a common goal, such as herd hunting), desocialization (low values of lamed), and misbelief. We can also see that the behaviour of an individual might lead or mislead the behaviour of his whole society. These emergent behaviours were not designed in I&I, but emerge from the properties of the environment and interactions of the animats.
The developed experiments tried to show and illustrate the capabilities of our BVL, the intelligence and emergent cognition in BeCA, and the emergence of social phenomena in I&I. Since we have obtained unexpected results in our experiments, showing emergent properties also present in natural systems, we believe that further experimentation (probably also carried out by motivated readers) would lead to the discovery of more emergent properties, and suggestions for refining our models.
Also, our present computing capabilities limit us to societies of less than ten individuals. Experiments with a higher number of animats would also bring interesting results and understandings.
1. Yabloko is apple, and koshki are cats.
2. See Section 3.5.
3. See Section 3.6.
4. See Section 3.7.
5. See Section 3.8.1.
6. See also Section 3.8.1.
7. See Section 3.8.2.
8. Doran (1998) presents an extensive discussion on collective misbelief in artificial societies.
Carlos Gershenson